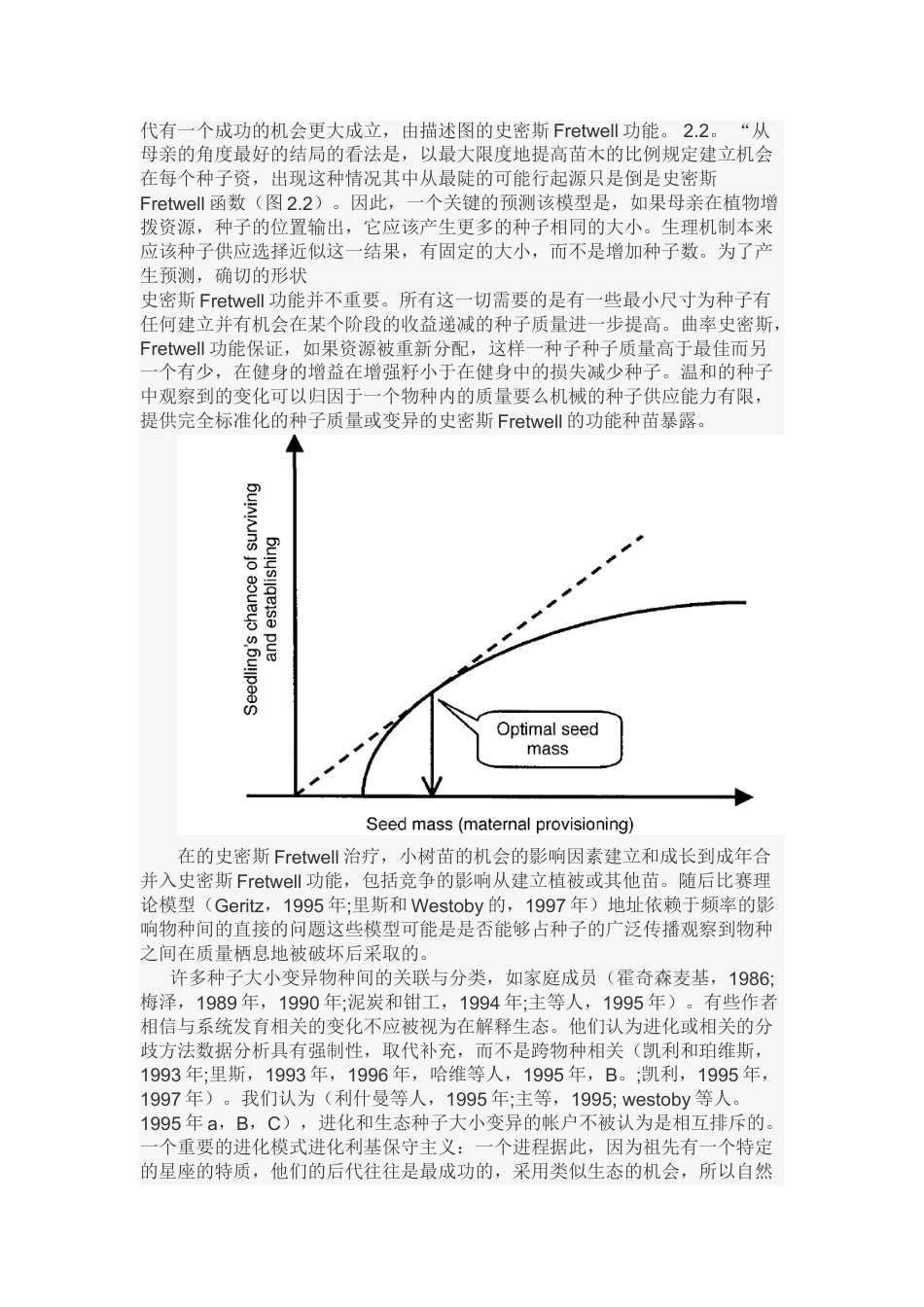
第二章进化生态学的种子大小MichelleR.Leishman,IanJ.Wright,AngelaT.MolesandMarkWestobyDepartmentofBiologicalSciences,MacquarieUniversity,Sydney,NewSouthWales,Australia简介在生态物种中,种子的质量是一个占有举足轻重的地位的特点。它是连接与营养生长的生态繁殖和幼苗建立的生态环境,是在很大程度上断开的其他方面的战略部门。(Grimeetal.,1988;Shipleyetal.,1989;LeishmanandWestoby,1992).在种子植物种类之总体之间间的世界,有一个惊人的多样性的形状和尺寸,范围从尘埃种子植物的种子和一些saprothytic寄生物种(大概10¯6克)。跨越是个数量级的Lodoicea和seychellarum两种椰子(104克)(Harperetal.,1970).。在物种内,种子的大小的跨越不到半个数量级(约四倍:Michaelsetal.,1988).。大多数物种的变异发生在植物中而不是植物与族群之间(Michaelsetal.,1988;Obeso,1993;VaughtonandRamsey,1998),这表明影响环境发育的不是母系的遗传差异。本章关注的是不同物种之间的种子大小,与群落变化和团体构成的结果。在过去的十到十五年间,在生态学的种子质量方面有了很大进展。不像其他领域的比较植物生态学,我们有从几个不同尺度和研究风格上的丰富的出版信息。以及对几个物种逐一进行了田间试验和数量研究,我们还对大量的物种进行了简单的实验(10到50),在不同的大陆量化种子质量及相关整个植物群落类型(上百个群落)和一致性测试。大规模量化开始早在Salisbury(1942)和Baker(1972),但是在过去的十年有了很大的增加和提高(e.g.Mazer,1989,1990;LeishmanandWestoby,1994a;Leishmanetal.,1995;ErikssonandJakobsson,1998).详细实验涉及的只有少数物种与大跨越物种数量的工作是相辅相成的,从一个特定的实验可以给出一个广泛的结果的强烈感觉。许多文献探讨如何自然选择种子大小可能会受到各种环境因素的影响。在这种情况下,乍看之下令人惊讶的是在不同的群落下种子的大小显示五到六个数量级(Leishmanetal.,1995;Fig.2.1).而且,完全不同的习性的种子的大小分部之间有强烈的重叠。在温带,物种间种子大小在群落之间的差异只占4%的变化(Leishmanetal.,1995)。在热带地区和温带地区的差异都有点大(Lordetal.,1995),但是一个栖息地的变化在物种变异的总体之间仍然是一个非常大的组成部分。这种可能形成广泛的物种变化的替代性机制还尚未完全制定的理论,暴露在强烈的假设性实验中要少得多。(图见原文)种子大小的组成和成分种子包括胚胎和胚乳(有时被称为种子储备),加上保护种子涂层或种皮。许多种子具有独特的分散种子附属物,如风传播的羽毛和毛发,挂钩和分散粘附的倒钩,油质体被蚁扩散和假种皮或肉被脊椎动物扩散。这些疏散附属物和种子,被称为水硬铝石。分散结构的质量种子质量的比例是种皮之间差异很大的物种。种子大小没有任何单个度量值是适合于所有目的。讨论幼苗建立,种子储备质量最能反映苗期可用的资源。讨论的对象有一个给定的扩散机制,种子质量包括种子大衣是最相关的移动大小。讨论成本,每种子生产的母系,整个水硬铝石的质量比种子质量较好,但仍然不是一个完整的措施再生产的一切费用。westoby(1998年)的建议干种子的质量作为生态战略轴,包括种皮,但不包括分散结构,部分作为替代措施之间的妥协,部分原因是因为它是最容易测量和部分保持着与大多数现有数据的可比性。同时,越来越多的研究去剖析组件的故障,它似乎与植物的传播体有关(如Westoby等,1990;胡拉多和Westoby,1992年,1994利什曼和Westoby的,C)。幸运的是,,数据集涵盖了广泛的种子质量,替代措施将密切相关.。假设,例如,这两个品种的种子质量的扩散结构质量在0到300%之间,这只能推翻相对种子质量排名的物种与种子传播体质量的排名,其种子的质量相差不到三个因素。因此,种子质量的规模设置跨越订单数据,记录干种子传播体的质量和干种子质量之间有很强的正相关关系,例如:在新南威尔士州西部,N=243,r2=0.71;澳大利亚中部,N=199,r2=0.83;悉尼,N=286,r2=0.97。另一方面,跨物种之间的仅仅(说)三到四倍,在种子质量,种子传播体质量排名可能与种子质量排名...